Tümör nekroz faktörü alfa
| TNF | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Tanımlayıcılar | |||||||||||||||||||||||||
| Başka adlar | TNF, tümör nekroz faktörü, tümör nekroz faktörü alfa, TNF-alfa, TNF-α, tümör nekrozu faktörü, tümör nekrozu faktörü alfa, tümör nekroz faktör, tümör nekroz faktör alfa, TNLG1F, DIF, TNFSF2, TNFA, tümör nekrozis faktör, tümör nekrozis faktör alfa | ||||||||||||||||||||||||
| Dış kimlikler | OMIM: 191160 MGI: 104798 HomoloGene: 496 GeneCards: TNF | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Ortologlar | |||||||||||||||||||||||||
| Türler | İnsan | Fare | |||||||||||||||||||||||
| Entrez |
|
| |||||||||||||||||||||||
| Ensembl |
|
| |||||||||||||||||||||||
| UniProt |
|
| |||||||||||||||||||||||
| RefSeq (mRNA) |
|
| |||||||||||||||||||||||
| RefSeq (protein) |
|
| |||||||||||||||||||||||
| Yerleşim (UCSC) | Krom. 6: 31.58 – 31.58 Mb | n/a | |||||||||||||||||||||||
| PubMed araması | [2] | [3] | |||||||||||||||||||||||
| Vikiveri | |||||||||||||||||||||||||
| |||||||||||||||||||||||||




Tümör nekroz faktörü (TNF, kaşeksin veya kaşektin; önceleri tümör nekroz faktörü alfa veya TNF-α olarak adlandırılıyordu); sistemik inflamasyonda yer alan bir hücre sinyal proteinidir (sitokindir) ve akut faz reaksiyonunu oluşturan sitokinlerden biridir. CD4+ lenfositler, NK hücreleri, nötrofiller, mast hücreleri, eozinofiller ve nöronlar gibi diğer birçok hücre tipi tarafından üretilebilmesine karşın, esas olarak aktif makrofajlarca üretilir.[4] TNF, homolog bir TNF alanına sahip çeşitli transmembran proteinlerden oluşan TNF süper ailesinin bir üyesidir.
TNF'nin birincil rolü bağışıklık hücrelerinin düzenlenmesidir. Bir endojen pirojen olan TNF; ateşi, apoptotik hücre ölümünü, kaşeksiyi, inflamasyonu tetikleyebilir, tümörijenezi ve viral replikasyonu (virüs çoğalmasını) baskılayabilir ve IL-1 ile IL-6 üreten hücreler yoluyla sepsise yanıt verebilir. TNF üretiminin bozukluğu; Alzheimer hastalığı,[5] kanser,[6] majör depresyon,[7] sedef hastalığı[8] ve enflamatuvar bağırsak hastalığı (İBH)[9] gibi çeşitli insan hastalıklarında rol oynamaktadır. Tartışmalı olmasına karşın depresyon ve İBH çalışmalarında güncel olarak artmış TNF düzeyleri ile ilişki kurulmuştur.[10][11] Rekombinant TNF, tasonermin INN'nin altında bir immünostimülan olarak kullanılmaktadır. TNF, malignitenin düzenlenmesinde ektopik olarak üretilebilir ve hem aşırı üretiminin ilişkili olduğu kanserlerde ve hem de sekonder hiperkalsemi nedeninde paratiroit hormonununa paralellik gösterir.
Keşif
Bağışıklık sisteminin in vivo anti-tümöral yanıtı kuramı, doktor William B. Coley tarafından farkına varıldı. 1968'de, Irvine, Kaliforniya Üniversitesi'nden Gale A Granger, lenfositler tarafından üretilen sitotoksik bir etken bildirdi ve buna lenfotoksin (LT) adını verdi.[12] Bu keşif için övgüyü, aynı etkinliği aynı ay içinde yayımlanmış art arda bir dizi makalede bildiren Yale Üniversitesi'nden Nancy H. Ruddle paylaşmaktadır.[13] Daha sonra 1975'te New York Memorial Sloan-Kettering Cancer Center'dan Lloyd J. Old, makrofajlarca üretilen başka bir sitotoksik faktör bildirdi ve buna tümör nekroz faktörü (TNF) adını verdi.[14] Her iki faktör, fare fibrosarkom L-929 hücrelerini öldürme yeteneklerine dayanarak tanımlanmıştır. Bu kavramlar, Old'un grubundan Elizabeth Carswell ile işbirliğinde Avustralya Ulusal Üniversitesi'nden Ian A. Clark sıtma hastalığı ve endotoksin zehirlenmesinde TNF'nin aşırı üretiminin gösterilmeye çalışıldığı dizileme (sekanslama) öncesi çağın verileriyle çalışıldığı 1981 yılındaki sistemik hastalıklara dek uzanmaktaydı.[15][16]
LT ve TNF'yi kodlayan cDNA'lar 1984'te[17] klonlanmıştır ve bunların benzer oldukları ortaya çıkmıştır. TNF'nin reseptörüne bağlanması ve LT ile yer değiştirmesi, iki faktör arasındaki fonksiyonel homolojiyi doğrulamıştır. TNF ve LT'nin ardışık ve işlevsel homolojisi, TNF'nin TNF-α olarak ve LT'nin TNFβ olarak yeniden adlandırılmasına yöneltmiştir. 1985 yılında Bruce A. Beutler ve Anthony Cerami, kaşektinin (kaşeksiyi tetikleyen bir hormon) aslında TNF olduğunu keşfettiler.[18] Daha sonra TNF'yi ölümcül endotoksin zehirlenmesinin bir aracısı olduğunu saptadılar.[19] Kevin J. Tracey ve Cerami, ölümcül septik şokta TNF'nin kilit aracı rolünü keşfettiler ve monoklonal anti-TNF antikorlarının terapötik etkilerini belirlediler.[20][21] Daha yakın zamanlarda, Mark Mattson Laboratuvarı'nda yapılan araştırmalar TNF'nin, Mn-SOD ve Bcl-2 ekspresyonunu tetikleyen transkripsiyon faktörü NF-kappaB'nin etkinleşmesini kapsayan bir mekanizma ile nöronların ölümünü/apoptozunu önleyebileceğini göstermiştir.
Gen
İnsan TNF geni 1985'te klonlanmıştır.[22] Kromozom 6p21.3 ile eşleşir, yaklaşık 3 kilobaz içerir ve 4 ekson içerir. Son ekson, lenfotoksin alfa (LTA, bir zamanlar TNF-β olarak adlandırılmıştır) ile benzerlik gösterir.[23] TNF'nin 3' translasyon olmayan bölgesi, AU açısından zengin öge içerir.
Yapı
TNF, öncelikle kararlı homotrimerlerde düzenlenmiş 233-amino asit-uzun tip II transmembran proteini olarak üretilir.[24][25] Membranla birleştirilmiş formdan, çözünür homotrimerik sitokin (sTNF), metalloproteaz TNF alfa dönüştürücü enzim (ADAM17 olarak da adlandırılır) tarafından proteolitik bölünme yoluyla salınır.[26] Çözünür 51 kDa trimerik sTNF, nanomolar aralığın altındaki konsantrasyonlarda çözünmeye ve dolayısıyla da biyoetkinliğini yitirme eğilimindedir. İnsan TNF'sinin salgılanan formu, üçgen bir piramit biçimini alır ve yaklaşık 17-kDa ağırlığındadır. Hem salgılanan hem de zara bağlı formlar biyolojik olarak etkindir, ancak her birinin özgül işlevleri tartışmalıdır. Ancak, her iki formun da örtüşümleri ve farklı biyolojik etkinlikleri vardır.[27]
Ev faresi TNF'si ile insan TNF'si yapısal yönden farklıdır.[28] 17 kilodalton (kDa) TNF protomerleri (185 amino asit uzunluğunda); TNF ailesi için tipik bir 'jöle silindir' β-yapı oluşturmuş antiparalel β P-iplikleri ile birlikte iki antiparalel β kıvrımlı katmandan oluşmuştur; aynı zamanda viral kapsid proteinlerinde de bulunmuştur.
Hücre sinyallenmesi
TNF, iki reseptörü bağlayabilir: TNFR1 (TNF reseptörü tip 1; CD120a; p55/60) ve TNFR2 (TNF reseptörü tip 2; CD120b; p75/80). TNFR1 55-kDa ve TNFR2 75-kDa'dır.[29] TNFR1 çoğu dokuda dışa vurulur ve TNF'nin hem membrana bağlanmış hem de çözünür trimerik formları ile bütünüyle etkinleştirilebilirken, TNFR2 genel olarak bağışıklık sisteminin hücrelerinde bulunur ve TNF homotrimerinin membrana bağlı formuna yanıt verir. TNF sinyali ile ilgili çoğu bilgi TNFR1'den elde edildiğinden, TNFR2'nin rolü olasılıkla hafife alınmaktadır.
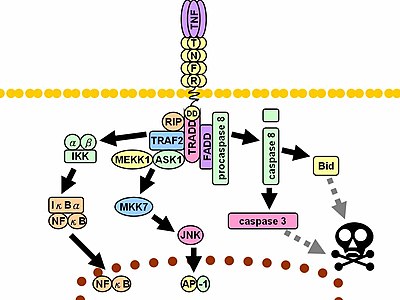
Kendi ligandlarıyla temasta TNF reseptörleri ayrıca, uçları TNF monomerleri arasında biçimlenen oluklara uyan trimerler oluşturur. Bu bağlanma, inhibitör protein SODD'nin hücre içi ölüm alanından ayrılmasına yol açan reseptörde üç boyutlu yapı değişikliğinin oluşmasına neden olur. Bu ayrışma, adaptör protein TRADD'nin, sonraki protein bağlanması için bir platform görevi gören ölüm alanına bağlanmasını sağlar. TRADD'nin bağlanmasını izleyen üç yol başlatılabilir.[30][31]
- NF-κB'nin etkinleştirilmesi: TRADD, TRAF2 ve RIP'yi devreye sokar. TRAF2 de çok bileşenli protein kinaz IKK'yi devreye sokar ve serin-treonin kinaz RIP'in bunu etkinleştirmesini sağlar. Normalde NF-κB'ye bağlanan ve yer değişimini inhibe eden inhibitör bir protein olan IκBα, IKK tarafından fosforile edilir ve daha sonra NF-κB salarak parçalanır. NF-κB, çekirdeğe yerleşen ve hücre sağ kalımını ve proliferasyonunu, enflamatuvar yanıt ve anti-apoptotik faktörlerde yer alan çok sayıda proteinin transkripsiyonuna aracılık eden heterodimerik bir transkripsiyon faktörüdür.
- MAPK yollarının etkinleştirilmesi: Üç ana MAPK kaskadından, TNF; stresle ilişkili JNK grubunun güçlü bir biçimde etkinleşmesini tetikler, p38-MAPK'nin orta düzeyde yanıtına neden olur ve klasik ERK'lerin en az düzeyde etkinleşmesinden sorumludur. TRAF2/Rac, MLK2/MLK3, TAK1, MEKK1 ve ASK1'in JNK uyarıcı ters yönlü kinazlarını etkinleştirir (ya doğrudan ya da sırasıyla GCK'ler ve Trx yoluyla). SRC-Vav-Rac ekseni MLK2/MLK3'ü etkinleştirir ve bu kinazlar daha sonra JNK'yi etkinleştirirken MKK7'yi fosforiller. JNK çekirdeğe taşınır, c-Jun ve ATF2 gibi transkripsiyon faktörlerini etkinleştirir. JNK yolağı hücre farklılaşmasını, proliferasyonunu kapsar ve genellikle proapoptotiktir.
- Ölüm sinyalinin tetiklenmesi: TNFR süper ailesinin tüm ölüm etki alanı içeren üyeleri gibi TNFR1 de ölüm sinyallenmesinde yer alır.[32] Bununla birlikte, TNF'nin neden olduğu hücre ölümü, enflamatuvar süreçteki baskın işlevleriyle karşılaştırıldığında oldukça küçük bir rolü vardır. Ölüme neden olan yeteneği diğer aile üyelerine (Fas gibi) oranla zayıftır ve genellikle NF-κB'nin anti-apoptotik etkileri ile maskelenir. Bununla birlikte TRADD, FADD'ı bağlarken daha sonra sistein proteaz kaspaz 8'i devreye sokar. Yüksek bir kaspaz-8 yoğunluğu, otoproteolitik etkinleşmesini ve sonrasında etkileyici kaspazların yarılmasını tetikleyerek hücre apoptozuna yol açar.
Yukarıdaki yolların aracılık ettiği sayısız ve sıklıkla çelişen etkiler, geniş çaplı çapraz iletişimin varlığını gösterir. Örneğin NF-KB; Cı-FLIP, Bcl-2 ve cIAP1/cIAP2, ölüm sinyallenmesinde müdahale eden inhibitör proteinlerin transkripsiyonunu arttırır. Öte yandan, etkinleştirilmiş kaspazlar RIP, IKK ve NF-κB'nin alt birimleri de içinde olmak üzere NF-κB yolunun çeşitli bileşenlerini ayırır. Hücre tipi, diğer sitokinlerin eş zamanlı uyarılmasını veya reaktif oksijen türlerinin (ROS) miktarı gibi başka etkenler dengeyi bir yol veya diğeri lehine değiştirebilir.[kaynak belirtilmeli] Böyle karmaşık sinyallenme bunu sağlama altına alır, TNF salındığında çok farklı işlevlere ve koşullara sahip olan çeşitli hücrelerin inflamasyona uygun biçimde yanıt verebilmesini sağlar.[kaynak belirtilmeli] Hayvan modellerinde TNF, seçici olarak otoreaktif T hücrelerini öldürür.[33]
Farelerde yapılan çalışmalarda TNF'nin sınıf 1 MHC (major histokompabilite kompleksi, MHK) antijenlerinin fare astrositlerindeki ekspresyonunu tetiklediği gösterilmiştir.[34] Sıçanlarda yapılan başka bir çalışmada TNF'nin sıçan nöronlarında TNFR1 üzerinden sınıf 1 MHC'nin ekspresyonunu tetiklediği gösterilmiştir.[35] İnsan iskelet kaslarında sınıf 2 MHC'lerin ekspresyonunu düzenlediği gösterilmiştir.[36]
Enzim düzenlenmesi
Bu protein allosterik düzenlenmenin morfein modelini kullanabilir.[37]
Fizyoloji
TNF'nin önceleri yalnızca makrofajlar[38] tarafından üretildiği düşünülmekteydi; ancak lenfoid hücreler, mast hücreleri, endotelyal hücreler, kardiyak miyositler, yağ dokusu, fibroblastlar ve nöronlar da içinde olmak üzere çok çeşitli hücre tipleri tarafından da üretilebilir.[4] Lipopolisakkarit, diğer bakteriyel ürünler ve İnterlökin-1'e (IL-1) yanıt olarak büyük miktarlarda TNF salınır. Deride mast hücreleri, enflamatuvar uyaran (örn. LPS) üzerine salınabilen, önceden oluşturulmuş TNF'nin baskın kaynağı gibi görünmektedir.[39]
Genellikle IL-1 ve İnterlökin-6 (IL-6) ile birlikte çeşitli organ sistemleri üzerinde bir dizi etkisi vardır:
- Hipotalamusta :
- Kortikotropin salgılatıcı hormonun (CRH) salınmasını uyararak hipotalamik-hipofiz-adrenal eksenin uyarılması
- İştahı bastırmak
- Ateş
- Karaciğerde: C-reaktif protein ve diğer bazı aracılarda bir artışa yol açan akut faz yanıtını uyarmak. Ayrıca insülin sinyalini bozan insülin reseptör substratı-1'in (IRS-1) serin-fosforilasyonunu kolaylaştırarak insülin direncini tetikler.
- Nötrofiller için güçlü bir kemo-çekici (kemoatraktant) ve endotelyal hücreler üzerinde yapışma moleküllerinin ekspresyonunu kolaylaştırarak nötrofillerin göç etmesine yardımcı olur.
- Makrofajlarda: Fagositozu ve IL-1 oksidanların ve inflamatuvar lipit Prostaglandin E2'nin (PGE 2) üretimini uyarır.
- Diğer dokularda: Artan insülin direnci. TNF, insülin reseptörü serin kalıntılarını fosforiller, sinyal iletimini engeller.
- Metabolizma ve besin alımı üzerinde: Acı tat algısını düzenler.[40]
TNF konsantrasyonundaki yerel bir artış, enflamasyonun ana belirtilerinin ortaya çıkmasına neden olacaktır: Isı, şişme, kızarıklık, ağrı ve işlev kaybı.
Yüksek TNF konsantrasyonları şok benzeri semptomlara neden olurken, düşük TNF konsantrasyonlarına uzun süre maruz kalmak, tüketen bir sendrom olan kaşeksiyle sonuçlanabilir. Bu durum örneğin kanser hastalarında bulunabilir.
Said ve ark., TNF'nin, PD-1'in PD-L ile bağlanması sonrasında monositler tarafından IL-10 üretimine yol açan monositler üzerinde PD-1 düzeylerini yukarı regüle ederek CD4 T hücresi genişlemesinin IL-10'a bağlı inhibisyonuna ve görev yapmasına neden olduğunu göstermişlerdir.[41]
Pedersen ve ark., sepsis yanıtında TNF artışının egzersize bağlı miyokin üretimi tarafından baskılandığını gösterir. Akut egzersizin gerçek bir anti-enflamatuvar yanıtı tetikleyip tetiklemediğini incelemek için, daha önce dinlenmeye veya egzersize rastgelelikle dağıtılan sağlıklı gönüllülere düşük dozda E. coli endotoksin uygulamasıyla bir tür 'düşük dereceli inflamasyon' modeli oluşturulmuştur. Dinlenen deneklerde endotoksin, dolaşımdaki TNF düzeylerinde 2 ila 3 kat artışa neden olmuştur. Buna karşılık, denekler 3 saat ergometre döngüsü gerçekleştirdiğinde ve 2.5 saatte endotoksin bolusu aldığında deneklerde TNF yanıtı bütünüyle baskılanmıştır.[42] Bu çalışma, akut egzersizin TNF üretimini engelleyebileceğine ilişkin bazı kanıtlar sunmaktadır.[43]
Farmakoloji
TNF, romatoid artrit, ankilozan spondilit, inflamatuvar bağırsak hastalığı, sedef hastalığı, hidradenit süpürativa ve refrakter astım gibi otoimmün bozukluklarla ilişkili klinik sorunların çoğuna neden olan enflamatuvar yanıtı kolaylaştırır. Bu bozukluklar bazen bir TNF inhibitörü kullanılarak tedavi edilir. Bu inhibisyon, adalimumab (Humira), sertolizumab pegol (Cimzia), doğrudan TNF'ye bağlanan infliksimab (Remicade) gibi bir monoklonal antikorla veya TNF'ye TNFR'den daha fazla ilgi ile bağlanan etanersept (Enbrel) gibi tuzak reseptör füzyon proteiniyle başarılabilir.[44]
Öte yandan TNF inhibitörleri ile tedavi edilen bazı hastalar, hastalıklarının şiddetlenmesine veya yeni otoimmünitenin başlamasına neden olur. TNF'nin de immünosüpresif bir yönü var gibi görünmektedir. Olası bir mekanizma için bir açıklama, TNF'nin Tümör nekroz faktörü reseptörü 2'ye (TNFR2) bağlanması nedeniyle, düzenleyici T hücreleri (Treg'ler) üzerinde olumlu bir etkiye sahip olduğudur.[45]
Anti-TNF tedavisi, kanser tedavisinde ancak ılımlı etkiler göstermiştir. Renal hücreli karsinomun infliksimab ile tedavisi, bazı hastalarda hastalığın uzun süreli dengesinin sağlanması ile sonuçlanmıştır. Etanersept, IL-6 ve CCL2'nin aşağı regülasyonu yoluyla bazı hastalarda hastalığın uzun süreli dengesinin sağlanmasını gösteren meme kanseri ve yumurtalık kanseri olan hastaların tedavisi için sınanmıştır. Öte yandan, ilerlemiş pankreas kanseri olan hastaların tedavisi için gemsitabine infliksimab veya etanersept eklenmesi, plasebo ile karşılaştırıldığında etkinlikteki farklılıklarla bir ilişkisi kurulamamıştır.[46]
Etkileşimler
TNF'nin TNFRSF1A ile etkileşime girdiği gösterilmiştir.[47][48]
Terminoloji
LTα artık TNFβ olarak anılmadığından dolayı[49] önceki gen sembolü olarak TNF-α, HGNC (HUGO Gen Adlandırma Komitesi) veritabanında gösterildiği gibi artık basitçe TNF olarak adlandırılmaktadır.
Kaynakça
- ^ a b c ENSG00000230108, ENSG00000223952, ENSG00000204490, ENSG00000228321, ENSG00000232810, ENSG00000228849, ENSG00000206439 GRCh38: Ensembl release 89: ENSG00000228978, ENSG00000230108, ENSG00000223952, ENSG00000204490, ENSG00000228321, ENSG00000232810, ENSG00000228849, ENSG00000206439 - Ensembl, May 2017
- ^ "İnsan PubMed Başvurusu:". National Center for Biotechnology Information, U.S. National Library of Medicine. Arşivlenmesi gereken bağlantıya sahip kaynak şablonu içeren maddeler (link)
- ^ "Fare PubMed Başvurusu:". National Center for Biotechnology Information, U.S. National Library of Medicine. Arşivlenmesi gereken bağlantıya sahip kaynak şablonu içeren maddeler (link)
- ^ a b 9218250
- ^ Swardfager W, Lanctôt K, Rothenburg L, Wong A, Cappell J, Herrmann N (2010). "A meta-analysis of cytokines in Alzheimer's disease". Biol Psychiatry. 68 (10). ss. 930-941. doi:10.1016/j.biopsych.2010.06.012. PMID 20692646.
- ^ Locksley RM, Killeen N, Lenardo MJ (2001). "The TNF and TNF receptor superfamilies: integrating mammalian biology". Cell. 104 (4). ss. 487-501. doi:10.1016/S0092-8674(01)00237-9. PMID 11239407.
- ^ Dowlati Y, Herrmann N, Swardfager W, Liu H, Sham L, Reim EK, Lanctôt KL (2010). "A meta-analysis of cytokines in major depression". Biol Psychiatry. 67 (5). ss. 446-457. doi:10.1016/j.biopsych.2009.09.033. PMID 20015486.
- ^ Victor FC, Gottlieb AB (2002). "TNF-alpha and apoptosis: implications for the pathogenesis and treatment of psoriasis". J Drugs Dermatol. 1 (3). ss. 264-75. PMID 12851985.
- ^ Brynskov J, Foegh P, Pedersen G, Ellervik C, Kirkegaard T, Bingham A, Saermark T (2002). "Tumour necrosis factor alpha converting enzyme (TACE) activity in the colonic mucosa of patients with inflammatory bowel disease". Gut. 51 (1). ss. 37-43. doi:10.1136/gut.51.1.37. PMC 1773288 $2. PMID 12077089.
- ^ Mikocka-Walus AA, Turnbull DA, Moulding NT, Wilson IG, Andrews JM, Holtmann GJ (2007). "Controversies surrounding the comorbidity of depression and anxiety in inflammatory bowel disease patients: a literature review". Inflammatory Bowel Diseases. 13 (2). ss. 225-234. doi:10.1002/ibd.20062. PMID 17206706.
- ^ Bobińska K, Gałecka E, Szemraj J, Gałecki P, Talarowska M (2017). "Is there a link between TNF gene expression and cognitive deficits in depression?". Acta Biochim. Pol. 64 (1). ss. 65-73. doi:10.18388/abp.2016_1276. PMID 27991935.
- ^ Kolb WP, Granger GA (1968). "Lymphocyte in vitro cytotoxicity: characterization of human lymphotoxin". Proc. Natl. Acad. Sci. U.S.A. 61 (4). ss. 1250-5. Bibcode:1968PNAS...61.1250K. doi:10.1073/pnas.61.4.1250. PMC 225248 $2. PMID 5249808.
- ^ Ruddle NH, Waksman BH (Aralık 1968). "Cytotoxicity mediated by soluble antigen and lymphocytes in delayed hypersensitivity. 3. Analysis of mechanism". J. Exp. Med. 128 (6). ss. 1267-79. doi:10.1084/jem.128.6.1267. PMC 2138574 $2. PMID 5693925.
- ^ Carswell EA, Old LJ, Kassel RL, Green S, Fiore N, Williamson B (1975). "An endotoxin-induced serum factor that causes necrosis of tumors". Proc. Natl. Acad. Sci. U.S.A. 72 (9). ss. 3666-70. Bibcode:1975PNAS...72.3666C. doi:10.1073/pnas.72.9.3666. PMC 433057 $2. PMID 1103152.
- ^ Clark IA, Virelizier JL, Carswell EA, Wood PR (Haziran 1981). "Possible importance of macrophage-derived mediators in acute malaria". Infect. Immun. 32 (3). ss. 1058-66. PMC 351558 $2. PMID 6166564.
- ^ Clark IA (Temmuz 1982). "Suggested importance of monokines in pathophysiology of endotoxin shock and malaria". Klin. Wochenschr. 60 (14). ss. 756-8. doi:10.1007/BF01716573. PMID 6181289.
- ^ Pennica D, Nedwin GE, Hayflick JS, Seeburg PH, Derynck R, Palladino MA, Kohr WJ, Aggarwal BB, Goeddel DV (1984). "Human tumour necrosis factor: precursor structure, expression and homology to lymphotoxin". Nature. 312 (5996). ss. 724-9. Bibcode:1984Natur.312..724P. doi:10.1038/312724a0. PMID 6392892.
- ^ Beutler B, Greenwald D, Hulmes JD, Chang M, Pan YC, Mathison J, Ulevitch R, Cerami A (1985). "Identity of tumour necrosis factor and the macrophage-secreted factor cachectin". Nature. 316 (6028). ss. 552-4. Bibcode:1985Natur.316..552B. doi:10.1038/316552a0. PMID 2993897.
- ^ Beutler B, Milsark IW, Cerami AC (Ağustos 1985). "Passive immunization against cachectin/tumor necrosis factor protects mice from lethal effect of endotoxin". Science. 229 (4716). ss. 869-71. Bibcode:1985Sci...229..869B. doi:10.1126/science.3895437. PMID 3895437.
- ^ Tracey KJ, Beutler B, Lowry SF, Merryweather J, Wolpe S, Milsark IW, Hariri RJ, Fahey TJ, Zentella A, Albert JD (Ekim 1986). "Shock and tissue injury induced by recombinant human cachectin". Science. 234 (4775). ss. 470-74. Bibcode:1986Sci...234..470T. doi:10.1126/science.3764421. PMID 3764421.
- ^ Tracey KJ, Fong Y, Hesse DG, Manogue KR, Lee AT, Kuo GC, Lowry SF, Cerami A (Aralık 1987). "Anti-cachectin/TNF monoclonal antibodies prevent septic shock during lethal bacteraemia". Nature. 330 (6149). ss. 662-64. Bibcode:1987Natur.330..662T. doi:10.1038/330662a0. PMID 3317066.
- ^ Old LJ (1985). "Tumor necrosis factor (TNF)". Science. 230 (4726). ss. 630-2. Bibcode:1985Sci...230..630O. doi:10.1126/science.2413547. PMID 2413547.
- ^ Nedwin GE, Naylor SL, Sakaguchi AY, Smith D, Jarrett-Nedwin J, Pennica D, Goeddel DV, Gray PW (1985). "Human lymphotoxin and tumor necrosis factor genes: structure, homology and chromosomal localization". Nucleic Acids Res. 13 (17). ss. 6361-73. doi:10.1093/nar/13.17.6361. PMC 321958 $2. PMID 2995927.
- ^ Kriegler M, Perez C, DeFay K, Albert I, Lu SD (1988). "A novel form of TNF/cachectin is a cell surface cytotoxic transmembrane protein: ramifications for the complex physiology of TNF". Cell. 53 (1). ss. 45-53. doi:10.1016/0092-8674(88)90486-2. PMID 3349526.
- ^ Tang P, Klostergaard J (1996). "Human pro-tumor necrosis factor is a homotrimer". Biochemistry. 35 (25). ss. 8216-25. doi:10.1021/bi952182t. PMID 8679576.
- ^ Black RA, Rauch CT, Kozlosky CJ, Peschon JJ, Slack JL, Wolfson MF, Castner BJ, Stocking KL, Reddy P, Srinivasan S, Nelson N, Boiani N, Schooley KA, Gerhart M, Davis R, Fitzner JN, Johnson RS, Paxton RJ, March CJ, Cerretti DP (1997). "A metalloproteinase disintegrin that releases tumour-necrosis factor-alpha from cells". Nature. 385 (6618). ss. 729-33. Bibcode:1997Natur.385..729B. doi:10.1038/385729a0. PMID 9034190.
- ^ Palladino MA, Bahjat FR, Theodorakis EA, Moldawer LL (Eylül 2003). "Anti-TNF-α therapies: the next generation". Nature Reviews Drug Discovery. 2 (9). ss. 736-46. Bibcode:2003Natur.002..736B. doi:10.1038/nrd1175. PMID 12951580.
- ^ Olszewski MB, Groot AJ, Dastych J, Knol EF (Mayıs 2007). "TNF trafficking to human mast cell granules: mature chain-dependent endocytosis". J. Immunol. 178 (9). ss. 5701-9. doi:10.4049/jimmunol.178.9.5701. PMID 17442953.
In human cells, contrary to results previously obtained in a rodent model, TNF seems not to be glycosylated and, thus, trafficking is carbohydrate independent. In an effort to localize the amino acid motif responsible for granule targeting, we constructed additional fusion proteins and analyzed their trafficking, concluding that granule-targeting sequences are localized in the mature chain of TNF and that the cytoplasmic tail is expendable for endocytotic sorting of this cytokine, thus excluding direct interactions with intracellular adaptor proteins
- ^ Theiss. A. L. et al. 2005. Tumor necrosis factor (TNF) alpha increases collagen accumulation and proliferation in intestinal myofibrobasts via TNF Receptor 2. The Journal of Biological Chemistry. [Online] 2005. Available at: http://www.jbc.org/content/280/43/36099.long 18 Ocak 2020 tarihinde Wayback Machine sitesinde arşivlendi. Accessed: 21/10/14
- ^ Wajant H, Pfizenmaier K, Scheurich P (2003). "Tumor necrosis factor signaling". Cell Death Differ. 10 (1). ss. 45-65. doi:10.1038/sj.cdd.4401189. PMID 12655295.
- ^ Chen G, Goeddel DV (2002). "TNF-R1 signaling: a beautiful pathway". Science. 296 (5573). ss. 1634-5. Bibcode:2002Sci...296.1634C. doi:10.1126/science.1071924. PMID 12040173.
- ^ Gaur U, Aggarwal BB (2003). "Regulation of proliferation, survival and apoptosis by members of the TNF superfamily". Biochem. Pharmacol. 66 (8). ss. 1403-8. doi:10.1016/S0006-2952(03)00490-8. PMID 14555214.
- ^ Ban L, Zhang J, Wang L, Kuhtreiber W, Burger D, Faustman DL (2008). "Selective death of autoreactive T cells in human diabetes by TNF or TNF receptor 2 agonism". PNAS. 105 (36). ss. 13644-13649. doi:10.1073/pnas.0803429105. PMC 2533243 $2. PMID 18755894.
- ^ E. Lavi; A. Suzumura; D. M. Murasko; E. M. Murray; D. H. Silberger; S. R. Weiss (1988). "Tumor necrosis factor induces expression of MHC class I antigens on mouse astrocytes". Journal of Neuroimmunology. 18 (3). ss. 245-253. doi:10.1016/0165-5728(88)90102-6.
- ^ H. Neumann; H. Schmidt; A. Cavalié; D. Jenne; H. Wekerle (1997). "Major Histocompatibility Complex (MHC) Class I Gene Expression in Single Neurons of the Central Nervous System: Differential Regulation by Interferon (IFN)-γ and Tumor Necrosis Factor (TNF)-α". Journal of Experimental Medicine. 185 (2). ss. 305-316. doi:10.1084/jem.185.2.305. PMC 2196130 $2. PMID 9016879. KB1 bakım: PMC biçimi (link)
- ^ Christian W. Keller; Claudia Fokken; Stuart G. Turville; Anna Lünemann; Jens Schmidt; Christian Münz; Jan D. Lünemann (2011). "TNF-α Induces Macroautophagy and Regulates MHC Class II Expression in Human Skeletal Muscle Cells". Journal of Biological Chemistry. 286 (5). ss. 3970-3980. doi:10.1074/jbc.M110.159392. PMC 3030397 $2. PMID 20980264. 9 Mart 2021 tarihinde kaynağından arşivlendi. Erişim tarihi: 17 Mart 2020. KB1 bakım: PMC biçimi (link)
- ^ Selwood T, Jaffe EK (2011). "Dynamic dissociating homo-oligomers and the control of protein function". Arch. Biochem. Biophys. 519 (2). ss. 131-43. doi:10.1016/j.abb.2011.11.020. PMC 3298769 $2. PMID 22182754.
- ^ Olszewski, ve diğerleri. (2007). "TNF Trafficking to Human Mast Cell Granules: Mature Chain-Dependent Endocytosis". The Journal of Immunology. 178 (9). ss. 5701-5709. doi:10.4049/jimmunol.178.9.5701. PMID 17442953.
- ^ Walsh LJ, Trinchieri G, Waldorf HA, Whitaker D, Murphy GF (Mayıs 1991). "Human dermal mast cells contain and release tumor necrosis factor alpha, which induces endothelial leukocyte adhesion molecule 1". Proc. Natl. Acad. Sci. U.S.A. 88 (10). ss. 4220-4. Bibcode:1991PNAS...88.4220W. doi:10.1073/pnas.88.10.4220. PMC 51630 $2. PMID 1709737.
- ^ Feng P, Jyotaki M, Kim A, Chai J, Simon N, Zhou M, Bachmanov AA, Huang L, Wang H (Ekim 2015). "Regulation of bitter taste responses by tumor necrosis factor". Brain, Behavior, and Immunity. Cilt 49. ss. 32-42. doi:10.1016/j.bbi.2015.04.001. PMC 4567432 $2. PMID 25911043.
- ^ Said EA, Dupuy FP, Trautmann L, Zhang Y, Shi Y, El-Far M, Hill BJ, Noto A, Ancuta P, Peretz Y, Fonseca SG, Van Grevenynghe J, Boulassel MR, Bruneau J, Shoukry NH, Routy JP, Douek DC, Haddad EK, Sekaly RP (Nisan 2010). "Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection". Nat. Med. 16 (4). ss. 452-9. doi:10.1038/nm.2106. PMC 4229134 $2. PMID 20208540.
- ^ Starkie R, Ostrowski SR, Jauffred S, Febbraio M, Pedersen BK (2003). "Exercise and IL-6 infusion inhibit endotoxin-induced TNF-α production in humans". FASEB J. 17 (8). ss. 884-886. doi:10.1096/fj.02-0670fje. PMID 12626436.
- ^ Pedersen BK (Aralık 2009). "The diseasome of physical inactivity – and the role of myokines in muscle–fat cross talk". J Physiol. 587 (23). ss. 5559-5568. doi:10.1113/jphysiol.2009.179515. PMC 2805368 $2. PMID 19752112.
- ^ Haraoui, B; Bykerk, V (2007). "Etanercept in the treatment of rheumatoid arthritis". Therapeutics and Clinical Risk Management. 3 (1). ss. 99-105. doi:10.2147/tcrm.2007.3.1.99. PMC 1936291 $2. PMID 18360618.
- ^ Salomon BL, Leclerc M, Tosello J, Ronin E, Piaggio E, Cohen JL (2018). "Tumor Necrosis Factor α and Regulatory T Cells in Oncoimmunology". Front. Immunol. Cilt 9. s. 444. doi:10.3389/fimmu.2018.00444. PMC 5857565 $2. PMID 29593717.
- ^ Korneev, KV; Atretkhany, KN; Drutskaya, MS; Grivennikov, SI; Kuprash, DV; Nedospasov, SA (Ocak 2017). "TLR-signaling and proinflammatory cytokines as drivers of tumorigenesis". Cytokine. Cilt 89. ss. 127-135. doi:10.1016/j.cyto.2016.01.021. PMID 26854213.
- ^ Bouwmeester T, Bauch A, Ruffner H, Angrand PO, Bergamini G, Croughton K, Cruciat C, Eberhard D, Gagneur J, Ghidelli S, Hopf C, Huhse B, Mangano R, Michon AM, Schirle M, Schlegl J, Schwab M, Stein MA, Bauer A, Casari G, Drewes G, Gavin AC, Jackson DB, Joberty G, Neubauer G, Rick J, Kuster B, Superti-Furga G (Şubat 2004). "A physical and functional map of the human TNF alpha/NF-kappa B signal transduction pathway". Nat. Cell Biol. 6 (2). ss. 97-105. doi:10.1038/ncb1086. PMID 14743216.
- ^ Micheau O, Tschopp J (Temmuz 2003). "Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes". Cell. 114 (2). ss. 181-90. doi:10.1016/S0092-8674(03)00521-X. PMID 12887920.
- ^ Clark IA (Haziran–Ağustos 2007). "How TNF was recognized as a key mechanism of disease". Cytokine Growth Factor Rev. 18 (3-4). ss. 335-343. doi:10.1016/j.cytogfr.2007.04.002. PMID 17493863.
Dış bağlantılar
- Medical Subject Headings Tumor+Necrosis+Factor-alpha
- PDBe-KB'de UniProt: P01375 (Tümör nekroz faktörü) için PDB'de bulunan bütün yapısal bilgilere bir genel bakış.










